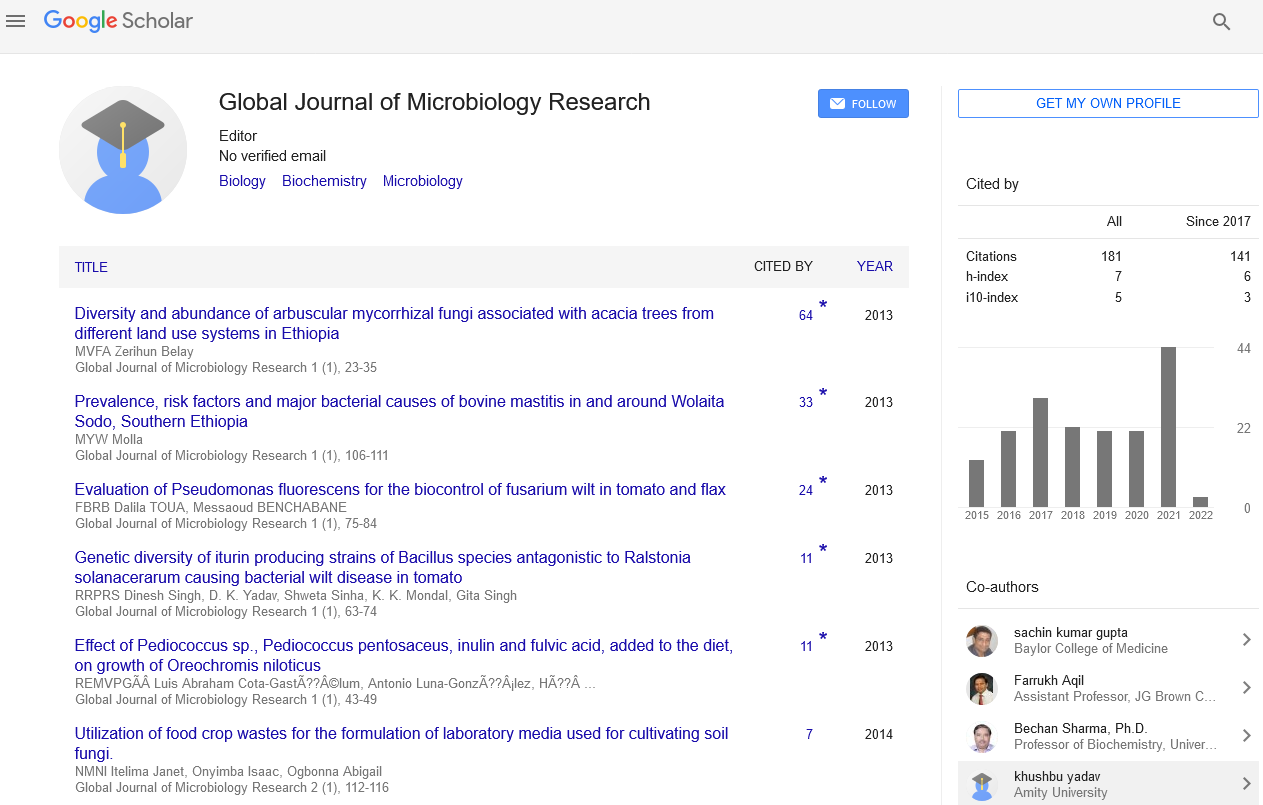
Global Journal of Microbiology Research received 161 citations as per Google Scholar report


Received: 04-Nov-2022, Manuscript No. GJMR-22-83184; Editor assigned: 07-Nov-2022, Pre QC No. GJMR-22-83184 (PQ); Reviewed: 21-Nov-2022, QC No. GJMR-22-83184; Revised: 29-Nov-2022, Manuscript No. GJMR-22-83184 (R); Published: 07-Dec-2022, DOI: 10.15651/GJMR.22.10.418
Intracellular pathogens face difficulties reproducing in a single host cell due to a lack of resources and space. We know how bacteria directly spread between host cells in cell culture, but we don't know how bacteria directly spread between host cells in vivo. Here, we report the identification of intracellular bacteria that spread between the intestinal epithelial cells of a naturally occurring host, the rhabditid nematode Oscheius tipulae, using filamentation. The bacteria, which are members of the newly discovered species Bordetella atropi, can infect the nematodes via the fecal-oral route and decrease the fecundity and lifespan of the hosts. UDP-glucose biosynthesis and sensing, a highly conserved pathway that other bacteria use to recognise rich environments and prevent cell division, are necessary for filamentation.
Our findings suggest that B. atropi triggers filamentation inside host cells using a pathway that typically controls bacterial cell size, facilitating cell-to-cell spread. Microbes frequently gain a significant advantage from the pathogenic invasion of host cells, such as a concentrated supply of metabolic resources and defence against extracellular immunity. However, because a host cell is a relatively small niche, intracellular pathogens find it advantageous to spread to other cells in order to make the most of the host's resources. All intracellular microbes can spread from the initial infected cell via extracellular escape, which makes them susceptible to host defences. However, some microbes have developed special invasion mechanisms that allow them to spread from an initial infected cell directly to uninfected host cells.
There are currently two main paradigms for how bacteria spread from cell to cell, and both of them centre on the manipulation of host actin by effectors. For cytoplasmic propulsion of the bacteria into the lateral membrane for uptake by neighbouring cells, Listeria monocytogenes, Shigella flexneri, and Rickettsia spp. can polymerize host actin in one mechanism. However, the invasion of neighbouring cells is made easier by the fusion of the lateral membranes in Burkholderia spp. Another method for the transfer of Ehrlichia chaffeensis involves the formation of actin-dependent filo podia on the surface of the target cell. It is unknown whether the pathogen is the one who initiates this method.
There may be additional mechanisms for cell-to-cell spreading and dissemination in vivo since none of these bacterial mechanisms have been directly observed outside of cell culture systems. Bacteria frequently alter the shape of their cells as a survival tactic to endure or thrive in a variety of environments. One example of this morphological plasticity is filamentation, which is frequently used as a survival tactic in toxic or demanding environments. When a bacterium divides longitudinally multiple times during filamentation, there is a noticeably larger increase in cell length and frequently no septation. The SOS response, which is triggered by environmental stressors like DNA damage and beta-lactam antibiotics, is known to cause filamentation in gram-negative bacteria. It has been noted that a number of bacterial pathogens, including H. influenzae, P. aeruginosa, and Y. pestis, form filaments in vivo, including inside host cells. To our knowledge, no intracellular pathogen, including bacteria or fungi, has ever been seen to use filamentation for cell-to-cell spreading.